A primeira lei de Mendel nos diz que os descendentes recebem um alelo do par de genes que determina uma característica.
Gregor Johan Mendel foi um monge agostiniano nascido no ano de 1822 que se interessou em explicar como as características dos pais são transmitidas a seus descendentes. Conhecido como o pai da genética, Mendel realizou todas as suas pesquisas sobre hereditariedade com ervilhas de cheiro (Pisum sativa), escolha que foi uma das razões de seu sucesso com suas pesquisas, pois essa leguminosa apresenta diversas vantagens como fácil cultivo, produção de grande quantidade de sementes, ciclo de vida curto, além de características contrastantes e de fácil identificação. Outro fato que contribuiu para o sucesso das pesquisas de Mendel foi que ele analisou apenas uma característica de cada vez, sem se preocupar com as demais características.
Em seus experimentos, Mendel teve o cuidado de utilizar apenas plantas de linhagens puras, por exemplo, plantas de sementes verdes que só originassem sementes verdes e plantas de sementes amarelas que só originassem sementes amarelas. Você deve estar se perguntando, como Mendel sabia que as plantas eram puras? Pois bem, para que ele tivesse certeza de qual planta era pura, ele as observava durante seis gerações, período de aproximadamente dois anos. Se durante essas gerações as plantas originassem indivíduos diferentes da planta inicial, elas não eram consideradas puras, mas se ocorresse o contrário e elas só originassem descendentes com as mesmas características da planta inicial, eram consideradas puras.
Uma vez constatado que as plantas eram puras, Mendel escolheu uma característica, por exemplo, plantas puras de sementes amarelas com plantas puras de sementes verdes, e realizou o cruzamento. Essa primeira geração foi chamada de geração parental ou geração P. Como resultado desse cruzamento, Mendel obteve todas as sementes de cor amarela e a essa geração denominou de geração F1. Os indivíduos obtidos nesse cruzamento foram chamados por Mendel de híbridos, pois eles descendiam de pais com características diferentes.
Em seguida, Mendel realizou uma autofecundação entre os indivíduos da geração F1,chamando essa segunda geração de geração F2. Como resultado dessa autofecundação, Mendel obteve três sementes amarelas e uma semente verde (3:1). A partir dos resultados obtidos, Mendel concluiu que como a cor verde não apareceu na geração F1, mas reapareceu na geração F2, as sementes verdes tinham um fator que era recessivo, enquanto as sementes amarelas tinham um fator dominante. Por esse motivo, Mendel chamou as sementes verdes de recessivas e as sementes amarelas de dominantes.
Em diversos outros experimentos, Mendel observou características diferentes na planta, como altura da planta, cor da flor, cor da casca da semente, e notou que em todas elas algumas características sempre se sobressaíam às outras.
→ Cada ser vivo é único e possui um par de genes para cada característica;
→ As características hereditárias são herdadas metade do pai e metade da mãe;
→ Os genes são transmitidos através dos genes;
→ Os descendentes herdarão apenas um gene de cada característica de seus pais, ou seja, para uma determinada característica, haverá apenas um gene do par, tanto da mãe quanto do pai.
Dessa forma, podemos enunciar a primeira lei de Mendel, também chamada de lei da segregação dos fatores da seguinte forma: “Todas as características de um indivíduo são determinadas por genes que se segregam, separam-se, durante a formação dos gametas, sendo que, assim, pai e mãe transmitem apenas um gene para seus descendentes”.
Gregor Mendel (1822-1884) foi um biólogo, botânico e monge austríaco. Descobriu as leis da genética, que mudaram o rumo da biologia.
Gregor Johann Mendel (1822-1884) nasceu em Heinzendorf, Áustria, no dia 22 de julho de 1822. Filho de camponeses, observava e estudava as plantas. Sua vocação científica desenvolveu-se paralela à vocação religiosa. Frequentou o ginásio de Troppau e estudou dois anos no instituto de Filosofia de Ormütz, depois Olomouc, hoje na República Tcheca.
Em 1843, com 21 anos, Mendel entrou para o Mosteiro Agostiniano de São Tomás, em Brünn, antigo Império Austro-Húngaro, hoje República Tcheca, onde foi ordenado padre e, passou a estudar teologia e línguas. Em 1847 ordenou-se e em 1851 foi enviado pelo abade à Universidade de Viena, para estudar ciências naturais, matemática e física. Três anos depois, voltou para Brünn.
Leis de Mendel
Gregor Mendel passou a dividir seu tempo entre lecionar numa escola técnica e plantar ervilhas-de-cheiro no jardins do mosteiro iniciando suas experiências com hibridação (cruzamento de espécies diferentes). Foram dez anos dedicados ao cruzamento de 22 variedades e acompanhando sete fatores com base na cor e forma da semente, forma da vagem, altura do caule etc., que lhe forneceram dados para formular as leis relativas à hereditariedade.
A primeira lei chamada “lei do monoibridismo”, foi resultado de uma série de cruzamentos com ervilhas durante gerações sucessivas e, mediante a observação do predomínio da cor (verde ou amarela), que lhe permitiu formular que existe nos híbridos uma característica dominante e uma recessiva. Cada caráter é condicionado por um par de fatores (genes) que se separam na formação dos gametas.
A segunda lei chamada “lei da recombinação ou da segregação independente” foi formulada com base na premissa segundo a qual a herança da cor era independente da herança da superfície da semente, ou seja, num cruzamento em que estejam envolvidos dois ou mais caracteres, os fatores que determinam cada um deles se separam de forma independente durante a formação dos gametas e se recombinam ao acaso, para formar todas as recombinações possíveis.
Reconhecimento Tardio
Os trabalhos de Mendel sobre hereditariedade que projetaram nova luz nas leis da herança não tiveram repercussão no meio científico da época. Sem estímulo para continuar e sobrecarregado com seus deveres administrativos no mosteiro, em 1868, ele abandonou por completo o trabalho científico. Sua obra permaneceu ignorada até o século XX, quando alguns botânicos, em pesquisas independentes, chegaram a resultados semelhantes e resgatam as leis de Mendel.
Johann Gregor Mendel faleceu em Brünn, República Tcheca, vítima de doença renal, no dia 6 de janeiro de 1884.
Obras de Gregor Mendel
Experiências Sobre Híbridos das Plantas (1865)
Alguns Híbridos do Hieracium Obtidos por Fecundação Artificial (1869)
Embriologia é o ramo da Biologia que estuda o desenvolvimento embrionário dos animais, isto é, as etapas pelas quais eles passam desde a fecundação até o nascimento.
A embriologia humana trata da gametogênese, ou seja, fertilização (união das células sexuais), desenvolvimento do embrião e da gravidez.
Até estar completamente formado o embrião passa por diversas etapas: segmentação, blastulação, gastrulação, neurulação, e organogênese.
FECUNDAÇÃO
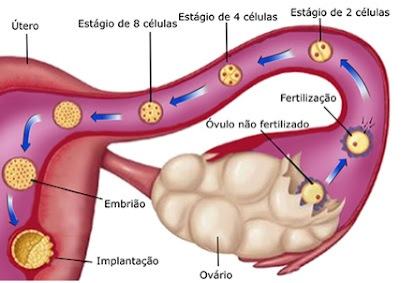
A Fecundação ou fertilização é o processo que ocorre quando os gametas masculinos e femininos encontram-se e o espermatozoide penetra o óvulo. Quando isto acontece, os nucléolos dessas células haploides (1n) fundem-se num só, formando a primeira célula diploide (2n) do novo ser vivo, o ovo ou zigoto.
Ao penetrar o óvulo, o espermatozoide perde seu flagelo e passa a ser chamado pronúcleo masculino.
A união dos pronúcleos masculinos e femininos chama-se cariogamia ou anfimixia (do grego amphi, dois, mixis, mistura).
A PRIMEIRA SEMANA
(SEGMENTAÇÃO ou CLIVAGEM DO ZIGOTO)
A clivagem consiste em repetidas divisões do zigoto, resultando em um rápido aumento do número de células. Primeiro, o zigoto se divide em duas células conhecidas como blastômeros; estas então se dividem em quatro blastômeros, oito blastômeros, e assim por diante.
A clivagem normalmente ocorre enquanto o zigoto atravessa a tuba uterina, rumo ao útero. O zigoto ainda se encontra contido pela substância gelatinosa muito espessa, a zona pelúcida, deste modo, ocorre um aumento no número de células sem que aumente a massa citoplasmática.
A divisão do zigoto em blastômeros começa cerca de 30 horas após a fertilização. Divisões subsequentes vão se seguindo e formam blastômeros progressivamente menores. Os blastômeros mudam de forma e se alinham, apertando-se uns contra os outros para formar uma esfera compacta de células conhecida como mórula. Este fenômeno, chamado de compactação, é provavelmente mediado por glicoproteínas de adesão da superfície celular.
A compactação permite uma maior interação célula-a-célula e constitui um pré-requisito para a segregação das células internas que formam o embrioblasto ou massa celular interna do blastocisto. A mórula (do latim, morus, amora), uma bola sólida de 12 ou mais blastômeros, é formada três dias após a fertilização e penetra no útero. Seu nome provém da sua semelhança com o fruto amoreira.
A SEGUNDA SEMANA
(FORMAÇÃO DA BLÁSTULA E IMPLANTAÇÃO DO BLASTOCISTO)
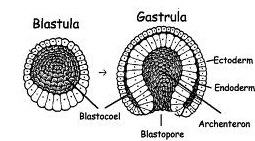
A blástula é o estágio de desenvolvimento embrionário em que, após sucessivas clivagens, centenas de células da mórula reorganizam-se agregadas e formam uma espécie de bola, com uma cavidade central repleta de líquido que denomina-se blastocele. Essas células formam uma camada celular chamada blastoderme.
A blástula sucede a mórula e antecede a gástrula. É, portanto, umas das primeiras fases de formação, antes que o embrião seja propriamente constituído.
Não se sabe exatamente quanto tempo o óvulo gasta para atravessar a trompa (oviduto). Presume-se que esse tempo seja de três a quatro dias. No sexto dia da fecundação, o blastocisto “fixa-se” no endométrio do útero, iniciando a fase de implantação. Nessa fase, o embrião vive à custa do material difusível através do endométrio, uma vez que suas reservas nutritivas (vitelo) são mínimas.
A implantação ocorre normalmente na parede posterior do corpo do útero, no espaço entre a abertura de glândulas do endométrio. Não é raro, porém, o blastocisto implantar-se em locais anormais, fora do corpo do útero. Em geral isso leva à morte do embrião, e a mãe sofre severa hemorragia durante o primeiro ou segundo mês de gestação.
SÚMULA DA IMPLANTAÇÃO
A implantação do blastocisto começa no fim da primeira semana e termina antes do final do segundo. O processo pode ser sumariado como se segue:
A zona pelúcida degenera (quinto dia). O desaparecimento da zona pelúcida resulta do aumento de tamanho do blastocisto e da degeneração causada por lise enzimática. As enzimas líticas são liberadas pelos acrossomos dos muitos espermatozoides que rodeiam e penetram parcialmente na zona pelúcida.
O blastocisto se liga ao epitélio endometrial (sexto dia).
O texofoblasto começa a se diferenciar em duas camadas, o suncicioblasto e o citotrofoblasto (sétimo dia).
O sincíciotrofiblasto evade os tecidos endometriais (capilares, glândulas, estroma) e o blastocisto começa a se implantar no endométrio (oitavo dia).
Aparecem no sinciciotrofoblasto lacunas repletas de sangue (novo dia)
O blastocisto penetra abaixo do epitélio endometrial
Redes lacunares são formadas pela fusão de lacunas adjacentes (décimo e décimo primeiro dias).
O sinciciotrofoblasto continua a erodir vasos sanguíneos endometriais, fazendo com que o sangue materno flua para fora das redes lacunares, e se estabelece, assim, uma circulação uteroplacentária primitiva. (décimo primeiro e décimo segundo dias)
A falha no epitélio endometrial desaparece gradualmente, enquanto o epitélio superficial se regenera. (décimo segundo e décimo terceiro dias).
Desenvolvem-se as vilosidades coriônicas primárias (décimo terceiro e décimo quarto dias)
A TERCEIRA SEMANA
Este é o inicio do período embrionário, que termina ao final da oitava semana. O rápido desenvolvimento do embrião a partir do disco embrionário, como resultado de numerosos eventos morfogenéticos, é caracterizado pela formação da linha primitiva, da notocorda e de três camadas germinativas a partir dos quais todos os tecidos e órgãos embrionários se desenvolvem.
NEURULAÇÃO
Aos processos envolvidos na formação da placa neural, das pregas neurais e no fechamento delas para formar o tubo neural dá-se o nome de neurulação. Estes processos estão completados pelo fim da quarta semana, quando ocorre o fechamento do neurósporo caudal. Durante a neurulação, o embrião pode ser chamado de neurula.
FORMAÇÃO DO TUBO NEURAL: A placa neural aparece como espaçamento do ectoderma embrionário localizado cefalicamente em relação ao nó primitivo. A placa neural é induzida a formar-se pela notocorda em desenvolvimento e pelo mesênquima adjacente. Um sulco neural, longitudinal, desenvolve-se na placa neural; o sulco neural ladeado pelas pregas neurais, que se juntam e se fundem para originar o tubo neural. O desenvolvimento da placa neural e seu dobramento para formar o tubo neural é chamado neurulação.
FORMAÇÃO DA CRISTA NEURAL: Com a fusão das pregas neurais para formar o tubo neural, células neuroctodérmicas migram ventrolateralmente para constituir a crista neural, entre o ectoderma superficial e o tubo neural. A crista neural logo se divide em duas massas que dão origem aos gânglios sensitivos dos nervos cranianos e espinhas. Outras células da crista neural migram do tubo neural e dão origem a varias estruturas.
DA QUARTA À OITAVA SEMANA
Essas cinco semanas constituem a maior parte do período embrionário, que se estende da terceira a oitava semana. Durante estas cinco semanas, que representam a maior parte do período embrionário, os principais órgãos e sistemas do corpo são formados a partir das três camadas germinativas.
No inicio da quarta semana, as dobras nos planos mediano e horizontal convertem o disco embrionário achatado em um embrião cilíndrico em forma de “C”. a formação das dobras cefálica caudal e laterais constitui uma sequência contínua de eventos que resultam numa constrição entre o embrião e o saco vitelino.
Durante o dobramento, a parte dorsal do saco vitelino é incorporado pelo embrião e dá origem ao intestino primitivo. Quando a região da cabeça se dobra ventralmente, parte do saco vitelino é incorporado pela cabeça embrionária em desenvolvimento como o intestino anterior. O dobramento da região da cabeça também faz com que a membrana orofaríngea e o coração sejam deslocados ventralmente, e que o encéfalo em desenvolvimento se transforme na parte mais cefálica do embrião.
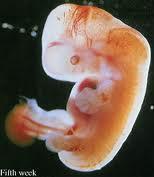
Quarta semana de gestação
Enquanto a região caudal se dobra ventralmente, uma parte do saco vitelino é incorporada à extremidade caudal do embrião, compondo o intestino posterior. A porção terminal do intestino posterior expande-se para constituir a cloaca. O dobramento da região caudal também resulta na membrana cloacal, na alantoide, e no deslocamento do pedículo do embrião para a superfície ventral dele.
O dobramento do embrião no plano horizontal incorpora parte do saco vitelino como intestino médio. O saco vitelino permanece ligado ao intestino médio por um estreito ducto vitelino. Durante o dobramento no plano horizontal, Formam-se os primórdios das paredes laterais e ventral do corpo.
Ao se expandir, o âmnio envolve o pedículo do embrião, o saco vitelino e a alantoide, compondo então um revestimento epitelial para nova estrutura chamada cordão umbilical.
As três camadas germinativas diferenciam-se em vários tecidos e órgãos, de modo que, ao final do período embrionário, estejam estabelecidos os primórdios dos principais sistemas de órgãos. A aparência externa do embrião é muito afetada pela formação do encéfalo, coração, fígado, somitos, membros, ouvidos, nariz e olhos. Com o desenvolvimento das estruturas, a aparência do embrião vai se alterando, e estas peculiaridades caracterizam o embrião como inquestionavelmente humano.
Com os primórdios das estruturas internas e externas essenciais se formam durante o período embrionário, a fase compreendia entre a quarta e a oitava semana constitui o período mais crítico do desenvolvimento. Distúrbios do desenvolvimento nesta altura podem originar grandes malformações congênitas no embrião.
DA NONA SEMANA ATÉ O NASCIMENTO
(O PERÍODO FETAL)
O período fetal, que começa nove semanas após a fertilização e termina com o nascimento, caracteriza-se pelo rápido crescimento corporal e diferenciação dos tecidos e órgãos. Uma mudança obvia é a diminuição relativa da velocidade de crescimento da cabeça, em comparação com o resto do corpo.
No início da vigésima semana aparece o lanugo e o cabelo, e a pele é recoberta pela vernix caseosa. As pálpebras permanecem fechadas na maior parte do período fetal, mas começam a se abrir por volta da vigésima sexta semana. Até então, o feto é usualmente incapaz de sobreviver fora do útero principalmente por causa da imaturidade do seu sistema respiratório.
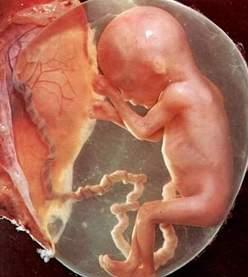
25ª semana de gestação
Até cerca da trigésima semana, o feto tem aparência avermelhada e enrugada por causa de sua pele fina e da relativa ausência de gordura subcutânea. Em geral, a gordura se forma rapidamente ao longo das últimas seis a oito semanas, dando ao feto um aspecto liso e rechonchudo. Esse período final (“de acabamento”) é dedicado principalmente à formação dos tecidos e à preparação dos sistemas envolvidos na transição do meio intra-uterino para o extrauterino, particularmente o sistema respiratório e cardiovascular. Fetos prematuros nascidos entre a vigésima sexta e a trigésima sexta semana em geral sobrevivem, mas fetos a termo têm maiores chances de sobrevivência.
As alterações que ocorrem no período fetal não são dramáticas quanto as do período embrionário, mas são muito importantes. O feto é menos vulnerável aos efeitos teratogênicos das drogas, vírus e radiação, mas estes fatores podem interferir com o desenvolvimento funcional normal, sobretudo do cérebro e dos olhos.
Existem várias técnicas disponíveis para avaliar as condições do feto e para diagnosticar certas moléstias antes do parto e anormalidades do desenvolvimento. Hoje em dia o médico pode determinar se um feto possui ou não certa doença ou uma malformação congênita, utilizando, por exemplo, a amniocentese e a ultrassonografia. O diagnóstico pré-natal pode ser realizado cedo o bastante para permitir o aborto seletivo de um feto defeituoso, se esta for a decisão da mãe e se o procedimento for legal; por exemplo quando forem diagnosticadas anomalias sérias, incompatíveis com a vida pós-natal.
Tipos de óvulos
Os ovos são classificados de acordo com a quantidade e distribuição do vitelo (reserva nutritiva), que garantem o desenvolvimento embrionário.
A partir desse componente de reserva, os óvulos podem ser:
ntral do ovo, em volta do núcleo. Ocorre nos artrópodes.
Tipos de óvulos
Os ovos são classificados de acordo com a quantidade e distribuição do vitelo (reserva nutritiva), que garantem o desenvolvimento embrionário.
A partir desse componente de reserva, os óvulos podem ser:
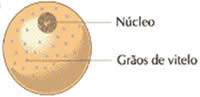
– Óvulos oligolécitos, isolécitos ou alécitos
Apresentam uma quantidade pequena de vitelo, que está distribuída de maneira uniforme pelo citoplasma. Ocorre nos espongiários, celenterados, equinodermas, protocordados e mamíferos.
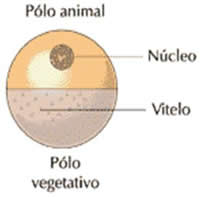
– Óvulos heterolécitos, mediolécitos ou telolécitos incompletos.
Apresenta uma quantidade média de vitelo que está concentrada abundantemente no pólo vegetativo. Ocorre nos platielmintes, moluscos, anelídeos e anfíbios.
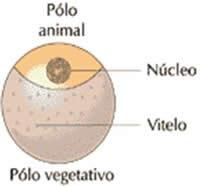
– Óvulos telolécitos completos ou megalécitos
Apresenta uma quantidade abundante de vitelo que ocupa praticamente todo o ovo, deixando o núcleo e o citoplasma numa pequena área do pólo animal, chamada cicatrícula ou disco germinativo.
Aparecem em cefalópodes, peixes, répteis e aves.
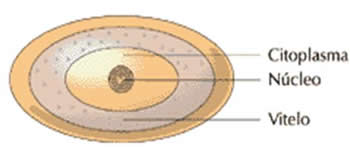
– Óvulos centrolécitos
O ovo possui uma abundante reserva de vitelo na região central do ovo, em volta do núcleo. Ocorre nos artrópodes.
Apresentam uma quantidade pequena de vitelo, que está distribuída de maneira uniforme pelo citoplasma. Ocorre nos espongiários, celenterados, equinodermas, protocordados e mamíferos.
– Óvulos heterolécitos, mediolécitos ou telolécitos incompletos.
Apresenta uma quantidade média de vitelo que está concentrada abundantemente no pólo vegetativo. Ocorre nos platielmintes, moluscos, anelídeos e anfíbios.
– Óvulos telolécitos completos ou megalécitos
Apresenta uma quantidade abundante de vitelo que ocupa praticamente todo o ovo, deixando o núcleo e o citoplasma numa pequena área do pólo animal, chamada cicatrícula ou disco germinativo.
Aparecem em cefalópodes, peixes, répteis e aves.
– Óvulos centrolécitos
O ovo possui uma abundante reserva de vitelo na região ce